Author note: at the publication date for this Insight, mouse-adapted SARS-CoV-2 viral strains were not available. However, two papers published after this Insight have demonstrated two different approaches for developing these mouse-adapted viral strains. Taconic has published a new Insight discussing this research.
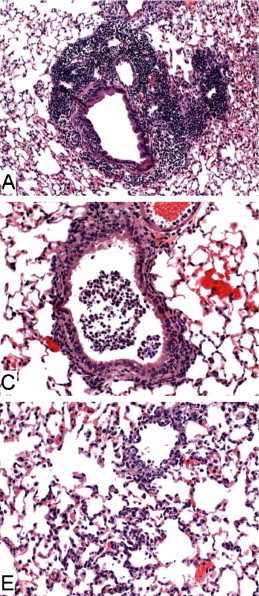
Figure Legend: Histopathology of lungs from aged 129S6 (A), B6 (C) and BALB/c (E) mice infected with SARS-CoV at 3 days post-infection. H&E stain. Adapted from Roberts, et al.5
Coronaviruses
Coronaviruses are a class of enveloped viruses that contain a positive-sense single-stranded RNA genome of roughly 30 kilobases in length1. In humans, these viruses can cause a variety of respiratory-related symptoms. Coronaviruses are responsible for 15-30% of all common cold cases, as well as more severe infections such as SARS, MERS, and COVID-192.Human coronaviruses were first characterized in patients with the common cold in the 1960s. Since then, multiple strains have been discovered, and there are now seven known strains that can infect humans: 229E, NL63, OC43, HKU1, MERS-CoV, SARS-CoV, and SARS-CoV-23. Of these coronaviruses, the SARS-CoV virus is the most closely related to the novel coronavirus, COVID-19 (SARS-CoV-2). Both viruses gain entry to human cells through the same cell surface receptor (ACE2) and contain similar genomic sequences. Thus, mouse models that were previously used to study the SARS-CoV infection may be valuable preclinical tools for studying the SARS-CoV-2 virus.
This Insight is part of our growing multi-part series on COVID-19. You may also be interested in:
- Mouse-adapted SARS-CoV-2 strains unlock broader mouse modeling of COVID-19
- Transgenic hACE2 mice for SARS-CoV-2 and COVID-19
- An Overview of Mouse Models for COVID-19
- Safety Testing Critical for COVID-19 Therapies and Vaccines
- Knockout Mice for the Study of Coronavirus Infections
Severe Acute Respiratory Syndrome (SARS)
SARS is the abbreviation for severe acute respiratory syndrome caused by the SARS-CoV coronavirus. SARS was first reported in early 2003 in Asia, and over the course of five months, spread to various Asian countries as well as to North America, South America, and Europe before the outbreak was contained. This virus was responsible for over 8,000 infections and 774 deaths4.Age-Related Mortality in SARS and COVID-19
It is well recognized that one of the consequences of aging is a decline in immune function known as immunosenescence. While both the adaptive and innate arms of the immune system are affected, the aging-associated deterioration of the adaptive immune system has been investigated in greater detail. This deterioration results in an impaired immune response to fighting new infections and developing immunity after vaccinations5. Additionally, the increased vulnerability of individuals aged 65 and older is well-established in influenza cases6.In both SARS and COVID-19 cases, the fatality rate increases with age7. The case fatality rate (CFR) is calculated by dividing the total number of deaths by the total number of cases, resulting in a percentage probability of death due to the infection. As shown in the Figure below, the lethality of COVID-19 increases steeply with age. In addition to immunosenescence, older individuals are more likely to have underlying health conditions that increase the fatality rate, such as diabetes mellitus, chronic lung disease, and cardiac disease.

Mouse models to study SARS
Since the SARS outbreak in 2003, multiple mouse models have been used to understand disease progression and pathogenesis. While no mouse model can replicate the aspects of this human disease entirely, there are several important aspects of viral infection that can be studied to inform treatment and vaccine development8,9. The SARS virus enters cells by binding to angiotensin-converting enzyme 2 (ACE2) and results in endocytosis of both the receptor and viral molecule10. This receptor is present on many cells, including arterial and venous endothelial cells, and abundantly present in epithelial cells in both the lungs and small intestine, potentially explaining the severe respiratory distress associated with this infection11,12.Multiple inbred strains have proven useful for SARS research, including BALB/c, C57BL/6, and 129S6. This Insight will focus on the use of aged mice specifically.
Aged BALB/c
When compared to young mice, aged BALB/c mice (12 to 14 months old) develop an enhanced and prolonged clinical illness following SARS infection. The SARS-CoV virus can replicate in the respiratory system in young BALB/c mice, but clinical symptoms of the virus are generally not evident13. Experiments in aged BALB/c mouse models have demonstrated that post-infection, proinflammatory cytokine levels are elevated, viral replication is persistent, and pneumonia develops in the lungs. This model is, therefore, instrumental in studying the age-related mortality seen in humans after SARS infection. One potential explanation for this is aging-associated immune senescence that has been described in both humans and mice14.Aged C57BL/6
Much like the BALB/c, aged C57BL/6 (B6) mice have used to study age-related mortality and alveolar damage seen in elderly patients with the SARS infection. In studies with clinical isolates of SARS-CoV, B6 mice displayed high levels of viral replication and proinflammatory chemokines in the lungs, as well as increased levels of SARS-CoV RNA in the brain, heart, liver, and spleen15. A serious complication associated with both SARS and COVID-19 is acute respiratory distress syndrome (ARDS). This condition is characterized by the activation of inflammatory cells and the subsequent release of inflammatory cytokines that contribute to lung injury and systemic inflammation16. B6 mice have been used to study the progression of ARDS as well as identify the biochemical and molecular pathways responsible for lung injury17. An added benefit of utilizing B6 mice for coronavirus research is the wide availability of genetically engineered mice (GEMs) on this background.Aged 129S6
Aged 129S6/SvEvTac mice have also been used in SARS studies, with weight loss and similar histopathological findings post-infection as in aged BALB/c and B6 mice8. According to Roberts, et al., when comparing experiments in young animals, "129S6 mice seem to be slightly more susceptible to disease associated with SARS-CoV infection than are BALB/c or B6 mice, as evidenced by an approximate 8% loss of body weight, with a nadir at days 5-7 [post-infection], and by the presence of mild interstitial pneumonitis at day 2 [post-infection]"8.Challenges in Using Aged Inbred Mice for COVID-19 Research
Aged inbred mouse models have been instrumental in understanding the disease pathogenesis of SARS-CoV and have produced highly reproducible data on viral replication and disease progression. However, inbred mouse strains cannot directly be put to use in COVID-19 studies just yet. SARS-CoV-2 uses the ACE2 protein to enter cells18. In vitro studies showed that ACE2 from many species facilitated entry into HeLa cells, but mouse ACE2 did not19. This poses a challenge for mouse studies of COVID-19, as clinical isolates may not be able to infect normal inbred mice. Strategies to address this challenge include the use of mouse-adapted SARS-CoV-2 viral strains (not yet reported to be available) or the use of mice that carry the human ACE2 gene. Many advancements can be expected in this rapidly-evolving field. Download the related Taconic Biosciences' White Paper:
Download the related Taconic Biosciences' White Paper: 
